Dolor persistente, aprendizaje neural y organización corporal
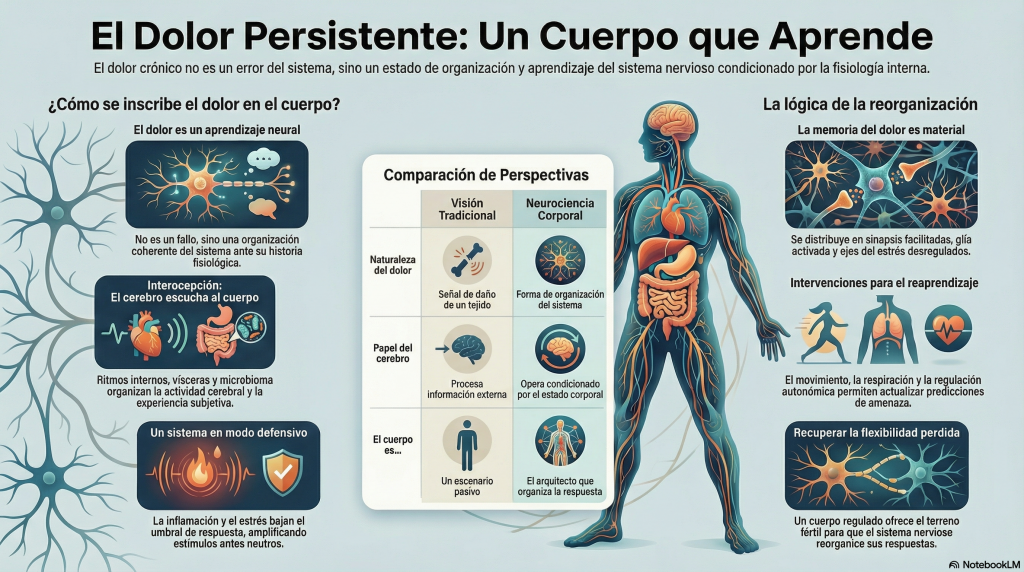
Cuando pienso el dolor persistente, no lo concibo como una señal prolongada ni como una lectura errónea del daño. Humildemente, creo que es una forma de organización del sistema nervioso.
Un estado que se ha aprendido. No como metáfora sino como consecuencia directa de la capacidad adaptativa del propio sistema.
Los mismos mecanismos que permiten aprender, anticipar y proteger —plasticidad sináptica, cambios duraderos de excitabilidad, reorganización de redes, activación glial— son los que, sostenidos en el tiempo, fijan el dolor como modo de funcionamiento.
El sistema no falla: responde con coherencia a la historia fisiológica que ha atravesado.
Desde ese lugar, el cuerpo nunca ha sido para mí un mero escenario donde el dolor acontece. Es el medio en el que ese aprendizaje se inscribe.
El trabajo desarrollado por Nazareth Castellanos, a la que tanto he escuchado y leído, ha permitido formular con una claridad especialmente fértil, algo que durante mucho tiempo permaneció implícito en la clínica: el cerebro no funciona desde la abstracción, sino desde un estado corporal previo.
No consulta al cuerpo como una fuente secundaria de información; opera condicionado por él. Respiración, señales viscerales, ritmo cardíaco, tono autonómico, microbioma y estado metabólico no acompañan la actividad cerebral: la organizan.

Esta afirmación no desplaza mi manera de pensar el dolor, la enriquece.
Si el cerebro opera siempre desde un estado corporal, entonces el dolor persistente no puede entenderse sin atender a ese estado. El mismo principio que explica la regulación explica también la cronificación.
El cuerpo no solo modula la actividad cerebral; cuando permanece demasiado tiempo en determinados registros fisiológicos, estrecha el repertorio posible del sistema nervioso y favorece aprendizajes rígidos.
Castellanos ha insistido en que el cerebro es profundamente sensible a la interocepción, a los ritmos internos, a la información que no llega del exterior sino del propio cuerpo. Desde ahí, la experiencia mental no se genera “en la cabeza”, sino en un diálogo continuo con lo visceral.
Interocepción: Capacidad del sistema nervioso para percibir, integrar e interpretar las señales internas del cuerpo. Incluye información procedente de las vísceras, el sistema cardiovascular, respiratorio, metabólico e inmunológico, y contribuye a la regulación del estado corporal, la emoción y la toma de decisiones. La interocepción no informa de lo que ocurre fuera, sino de cómo está el cuerpo, y desde ese estado condiciona la actividad cerebral y la experiencia subjetiva.
Pensado desde el dolor persistente, ese diálogo revela un desequilibrio: el cuerpo no solo informa, insiste. Y cuando esa insistencia adopta la forma de inflamación sostenida, estrés crónico o empobrecimiento de variabilidad fisiológica, termina escribiéndose como dolor.
El microbioma ocupa aquí un lugar que ya no puede considerarse marginal. En la neurociencia corporal, el eje intestino-cerebro permite comprender cómo lo periférico participa en la organización del estado mental y emocional.
En mi marco, ese mismo eje adquiere un peso específico adicional: el microbioma importa porque regula inflamación, permeabilidad intestinal y activación inmunitaria, y porque esas variables modifican directamente la excitabilidad del sistema nervioso. No introduce contenidos en la experiencia consciente, pero desplaza el punto desde el cual el sistema discrimina, aprende y anticipa. Antes de que el dolor sea vivido, el terreno ya se ha inclinado.
Ese aprendizaje comienza en lugares que no suelen aparecer en el relato subjetivo.
Los nociceptores no responden en el vacío; responden desde un entorno corporal concreto. Cuando ese entorno está marcado por inflamación sostenida, estrés prolongado o alteraciones metabólicas, los canales iónicos que regulan la transducción cambian su comportamiento.
El umbral baja, la respuesta se amplifica y estímulos antes neutros adquieren valor de amenaza. El cuerpo, pensado como organizador del estado —tal como propone Castellanos—, se convierte aquí en arquitecto del disparo nociceptivo.
La señal que emerge de ese contexto no viaja por un sistema neutral; los axones, inmersos en ese mismo estado corporal, pueden volverse eléctricamente inestables, generar actividad espontánea y amplificar descargas mínimas. El sistema ya no necesita estímulo externo para activarse: ha aprendido a anticipar desde dentro, a partir de su propia fisiología.
Cuando esa información alcanza la médula espinal, se integra, se refuerza, se aprende. La repetición facilita la transmisión, debilita la inhibición y consolida patrones de respuesta que dejan de depender del estado del tejido periférico. Esta reorganización no ocurre aislada del resto del organismo.
El estado corporal del que habla Castellanos —ese fondo autonómico, visceral y metabólico desde el que opera el cerebro— condiciona también qué señales ascienden, con qué intensidad y durante cuánto tiempo. La sensibilización central aparece entonces no como un fenómeno local, sino como la expresión medular de un cuerpo entero en modo defensivo.
En niveles superiores, el cerebro no espera a que algo ocurra para reaccionar. Predice. Anticipa. Interpreta desde señales internas que informan de cómo está el cuerpo.
Castellanos ha mostrado cómo respiración, interocepción y regulación autonómica influyen en esa dinámica predictiva. Cuando ese cuerpo ha permanecido demasiado tiempo en alerta, la predicción se rigidiza. El cerebro no se equivoca al doler: responde fielmente a un cuerpo que no ha podido dejar de avisar.
La glía desempeña aquí un papel decisivo. No como acompañante pasivo, sino como elemento que estabiliza el estado hiperexcitable y lo vuelve resistente al cambio.
La memoria del dolor no se guarda en un lugar concreto; se distribuye entre sinapsis facilitadas, axones inestables, glía activada y ejes neuroendocrinos desregulados. Es una memoria corporal, material, silenciosa, que no necesita relato para activarse. Y es también el lugar donde el cuerpo, en el sentido que propone Castellanos, se inscribe de forma duradera en el sistema nervioso.
Los sistemas hormonales refuerzan esta organización. La activación sostenida del eje del estrés pierde su capacidad reguladora y pasa a formar parte del problema. Aquí la convergencia es clara: lo que en la neurociencia del cuerpo aparece como regulación del estado, en el dolor persistente se manifiesta como su pérdida crónica, con consecuencias estructurales sobre excitabilidad, inflamación y plasticidad.
Desde esta mirada ampliada, las intervenciones corporales dejan de ser recursos blandos o complementarios.
Movimiento, reorganización sensoriomotora, regulación autonómica, atención a la respiración y a la interocepción actúan sobre distintos planos del mismo sistema. No compiten entre sí. Se necesitan. Un cuerpo más regulado —tal como lo piensa Castellanos— ofrece un terreno más fértil para que el sistema nervioso reaprenda; un sistema menos reactivo puede actualizar sus predicciones sin volver a activar la amenaza.
Integrar la neurociencia del cuerpo con una neurociencia del dolor persistente no suaviza el problema sino que lo vuelve más exigente. Obliga a pensar el dolor como el resultado de un aprendizaje distribuido, sostenido por un cuerpo que ha perdido flexibilidad y también a abandonar definitivamente las falsas dicotomías entre mente y cuerpo, entre señal y experiencia, entre biología y vivencia.
El cuerpo que organiza el cerebro es el mismo que, cuando permanece demasiado tiempo en alerta, fija el dolor como estado.
Pensar esta continuidad no promete alivio inmediato ni soluciones simples.
Permite algo más difícil pero más honesto: dejar de buscar el error en un punto aislado y empezar a escuchar la lógica completa de un sistema que ha aprendido bajo condiciones adversas sin culpabilizarlo, ni forzarlo a desaprender a golpes, sino para ofrecerle —con tiempo, cuidado y rigor— la posibilidad de reorganizarse.

Pensar así el dolor no consuela, pero define con precisión
Y a veces, nombrar bien es el primer acto de cuidado.